Modernos conocimientos sobre el origen de la vida
8.- Primera vida, ADN y el rol de las encimas
El paso de materia inerte hacia materia viva, se llevó a cabo sólo, cuando los Protobiontes no sólo entregan los procesos metabólicos, sino cuando estas células, también comenzaron a reproducirse basado en la información. La codificación de la información genética se produce por los ácidos nucleicos (más exacto: Con la ayuda del ácido desoxirribonucleico, ADN).
El desarrollo de los primeros ácidos nucleicos debió haber demorado muchos millones de años. Debido a que el ADN no puede reduplicarse y que las encimas, bióticamente no se forman ain el ADN, se presenta la pregunta, cómo se originó esta dependencia entre el ADN y las encimas en los primeros seres vivientes.
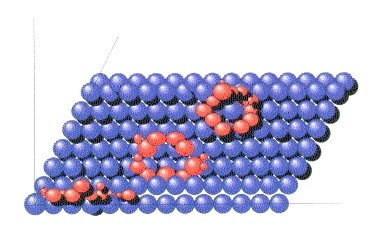
Un modelo esquemático de una toma de una microscopía de efecto túnel. El físico Heckl descubrió en la superficie de bisulfuro de molibdeno, extrañas estructuras redondeadas y también alargadas cuya distribución de densidad de electrones se destacaban notoriamente del trasfondo.
Un posible explicación dan los descubrimientos del físico Wolfgan Heckl , del grupo investigativo IBM de la universidad de Múnich. Heckl investigó diversos materiales portadores con propiedades dieléctricas en su idoneidad para la microscopía de efecto de túnel. El principio es el siguiente: Si se coloca una aguja, tan delgada como un átomo, sobre una superficie semiconductora muy liza, entonces comienza a fluir una corriente, si se aplica entra la aguja y la superficie una carga eléctrica. El tamaño de la corriente de túnel depende de la distancia hacia la superficie. Mediante esto es posible visualizar “valles” y “montañas” de capas atómicas individuales de la superficie del portador.
Heckl junto a diversos semiconductores sintéticos, también investigo cristales de bisulfuro de Molibdeno, el cual al igual que la pirita, también muestra propiedades conductoras. Cuando observaba en la pantalla de un computador las capas atómicas del bisulfuro, noto extrañas estructuras moleculares de forma de anillos con un diámetro de 4 nanómetros, como también estructuras alargadas, que parecían estar intercalados con el material portador. Estas estructuras moleculares sobre salen del plano del material portador y eran similares a las estructuras de la figura 9.
El material investigado por Heckl proviene del precámbrico, o sea tenía una edad de más de 650 millones de años. En un examen más detenido se descubrió, que estos anillos contenían carbono, por lo tanto debían ser de naturaleza orgánica. Heckl especula, que podría tratarse de anillos plásmidos, o sea de material genético de organismos de estructuras muy simples. Estos anillos plásmidos son representados por secuencias relativamente cortas de ADN que están adheridos en minerales. El descubrimiento sugiere la conclusión: El invento de los códigos genéticos, como también la estrecha unión entre encimas (que se componen de aminoácidos) y el AN (ácido nucleico) pudo haberse hecho en la superficie de sulfuros metálicos de este tipo. Con Heckl es posible pensar, que en la edad temprana de la Tierra, pudieron haber sido absorbidos aminoácidos y ácidos nucleicos, en varias capas sobre puestas, sobre sulfuros metálicos, y haberse formado así las primeras cadenas de t- ADN, que al “leer” y “traducir” están involucrados en el código genético en proteínas (respectivamente encimas).
9.- Cuasi especies e híper-ciclos
El profesor Manfred Eigen del instituto Max-Planck para química bio-física en Göttingen, persigue una teoría totalmente diferente. Según su punto de vista una determinada forma de ADN se desarrolló para formar una cadena de ARN, que tiene la capacidad de replicarse sin la ayuda de encimas.
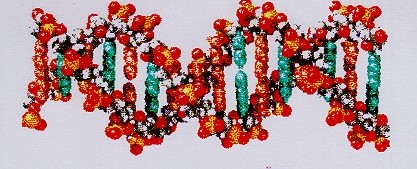
Recorte de una molécula de ADN. En el ADN dos cadenas de moléculas están enredadas entre sí en forma de espiral que son mantenidas juntas mediante bases nucleótidos. Las barras verticales azules y naranjas, representan aquí las bases nucleótidos; ellas simbolizan el código genético. La espiral se compone de un azúcar (desoxiribosa) que estpa químicamente atado con el pacido fosfórico. Existen 4 bases de nucleótidos. Adenina, Citosina, Guanina y Timina (en el ARN en vez de la Timina, el Uracilo). La secuencia (combinación) de estas bases en esta doble espiral, caracteriza el genoma del organismo presente y representa toda su genética y con esto las propiedades del ser. El ADN (aquí el ARN), es una bio-molécula fundamental. Durante la “lectura” del ADN, la doble hélice es desenvuelta en cadenas individuales y las secuencias de las bases son descifradas bioquímicamente secuencialmente. En ella está “escrita” que proteínas y encimas debe fabricar el organismo para permitir procesos metabólicos.
El “Híper-ciclo” fue postulado por EIGEN como primer sistema capaz de replicarse evolutivamente. En el híper-ciclo más simple se encuentran dos moléculas de ARN que en una interacción mutua que se originan de una solución de sustrato y se “reproducen”. Con esto se desacoplan dos o más cadenas de ARN (Ribocimas) auto reproductivas o Ribocimas y encimas que se acoplan para formar un auto-ciclo estable, que se mantiene así mismo y se replica (figura 11).
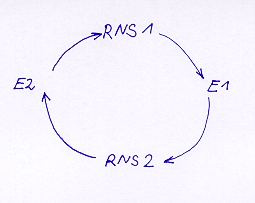
Aquí se muestra el principio esquemático de un llamado híper-ciclo, en el cual dos o más secuencias de ARN junto a las encimas (E) forman un ciclo y se mantiene mutuamente. Ambas secuencias depende entre si y no pueden permitirse expulsarse de esta carrera.
Eigen ideó el modelo del híper-ciclo, debido a que las grandes cuotas de errores en la proliferación de los poli nucleótidos (Ribosomas) que se auto, reproducen hacen que la información después de una determinado largo de secuencia se separan,. Después de cada paso de replicación pueden aparecer mutaciones, de manera que de una secuencia original se puede originar una “cola de cometa” de Ribosomas parecidos. Estos forman un conjunto de mutantes, las llamadas “cuasi especies”, que con respecto a la exactitud de copia, estabilidad y velocidad de replicación, compiten en1tre sí. Para poli nucleótidos, que se componen exclusivamente de pares de bases Guanina-Citosina , la tasa de error determinada experimentalmente es de aprox. un 1%, la cadena debería tener u largo máximo de 100 nucleótidos. Para poli nucleótidos, que se componen de pares de base Adenina-Uridina la tasa de error en la lectura en la replicación es unas diez veces del largo de la cadena, el largo de la cadena podría tener máximo 10 eslabones. Para garantizar una reproducción estable sin una disolución de la información, a lo largo de cualquier número de generaciones, es imposible concentrar un “genoma original” en una sola molécula.
Sin embargo si en una cuasi-especie se forman dos o más mutantes, que catalizan y estabilizan entre si su reproducción (eventualmente mediada por encimas), se forman sistemas cooperativas que se pueden reproducir por un largo período en forma estable y que frente a otros competidores en el conjunto de mutantes, poseen una determinante ventaja selectiva. Como condición debe valer, que cada ribosoma se componga de 50-100 eslabones de cadenas, para el “gen originario” se debe suponer una caudal guanina-Citosina de 50-100%, ya que las matrices ARN disponen de una relativamente reducida tasa de errores. De hecho Eigen y WINKLER-OSWATITSCH, han podido mostrar matemáticamente que las moléculas e-ARN recientes sugieren un estado originario, que correspondía exactamente a una distribución de casi-especies de moléculas auto reproducibles (EIGEN Y WINKLER-OSWATITSCH 1981). a mano de las secuencias variablea se pudo reconstruir matemáticamente la secuencia original hipotética (KÄMPFE 1992, pág. 201).
También es interesante, que los recientes t-ARNs muestran exactamente a las referidas propiedades (un alto componente de Guanina-Citosina de aprox. un 89% como un largo promedio de cadena de 76 nucleótidos) y por lo tanto tiene la misma proporción que matemáticamente se espera de la teoría, sin embargo si se supone una misteriosa “creatio ex nihilo” los descubrimientos quedan sin explicaciones.
10.- Conceptos afianzados en relación al origen de los primeros organismos (Protobiontes)
Los primero enfoque holísticos en relación al origen de la vida a base de ácidos nucleicos, fueron elaborados en 1972 por Kuhn y Kaplan y concretizados constantemente hasta hoy. De acuerdo a todo lo que sabemos hoy en día, se consideran dos hipótesis realistas, la “hipótesis de pasos múltiples” de Kuhn y la “hipótesis de aciertos múltiples” de Kaplan. En oposición a las teorías de los ácidos nucleicos de Kuhn, Kaplan y Eigen, que postulan, que oligonucleótidos, respectivamente poli nucleótidos fueron las macro moléculas primordiales de la Tierra, que se habrían formado sin Replicasas, respectivamente encimas, las hipótesis que se basan en la proteínas (Oparin), parten de la base, que primero se formaron los polipéptidos y las proteínas, que posteriormente, por vía catalítica formaron los ácidos nucleicos. En lo que sigue analizaremos y trataremos de atarlos con las hipótesis en de las proteínas con la teoría del híper-ciclo de Eigen
a). La hipótesis de los pasos múltiples:
En la teoría de Kuhn se basa la suposición, que las secuencias Oligonucleótidos y los ácidos nucleicos, se deben considerar como portadores primordiales de informaciones. Con esto es adecuada la hipótesis de las proteínas, para explicar la formación de estos ácidos nucleótidos a favor de este concepto hablan una serie de hallazgos:
Kaplan calculó, que la probabilidad de que una proteína pueda catalizar una reacción, se encuentra entre 10ˆ10 y 10ˆ14. También los Proteinoides funcionales, que en su estructura son bastante inconsistentes, muestran cinéticas del tipo enzimático, y mientras tanto se ha logrado evidenciar numerosos proteinoides de este tipo. Así por ejemplo se conocen Proteinoides abióticos con actividades ATPasa, Transaminasa, Esterasa, Catalasa y Peroxidasa, y hay que señalizar al sorprendente descubrimiento, que Proteinoides térmicos al agregarle una solución ATP construyen olgonucleótidos de la Adenina. En una solución acuosa a 90 °C se produce la poli condensación de TMP y dAMP bajo la acción catalizadora de histidina o, por ejemplo del polipéptido Poliornitina como lo han mostrada Oro y colaboradores. Se logró adherir a este tipo de oligonucleótidos, nucleótidos escalonadamente mediante condensación, de manera que al final se obtuvieron ácidos nucleicos más complejos. Incluso la síntesis controlada enzimáticamente, sin la intervención de matrices, de poli nucleótidos es conocida en el ejemplo de la Oβ-Replicasa ha sido mostrado en forma impresionante.
Estos y otros descubrimientos son un apoyo empírico, que justifica la suposición, tanto la encimas, como también ya los simples proteinoides funcionales (proto-encimas) pueden actuar como sintetasas (en parte también sin la necesidad de matrices). Los descubrimientos experimentales, además apoyan total y completamente la evaluación de Kaplan sobre la probabilidad de las proteínas (oides) funcionales. Sin embargo la actividad catalítica es bastante menor que la de las encimas bióticas, pero sin duda alguna hubiera bastado para posibilitar vida simple, ya que al comienzo de la evolución biótica aún no existían competidores de “mayor eficiencia”
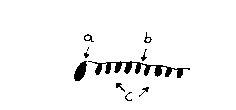
Modelo de nucleación (a) con cuerda recolectora (b) y moléculas de fijación (c). Según la teoría agregados enrollados estáticamente provenientes de la doble hélice (a), pudieron mostrar propiedades, oligonucleótidos lineales abiertos, atar las tal llamadas cuerdas colectoras, en cuyas bases de nucleótidos a su vez se pudieron unir brevemente Oligonucleótidos (Bien en general: Moléculas de fijación) por intermedio de puentes de Hidrógeno, que llevaron a la formación del t- ARN originario.
Oligonucleótidos simples posiblemente también pudieron haberse formado sin encimas en grandes cantidades, que se unieron en cuasi-especies híper cíclicamente organizados. Esta formación fue inicialmente al azar (divergente), pero como lo pudo demostrar, con el correr del tiempo voltearse a un proceso convergente que desembocó en una selección darwinista.
Para esto asumimos que se formaron aleatoriamente Oligonucleótidos, cuyas Ribosas todas son ópticamente destrogenas (Donde los Oligonucleótidos, con, por ejemplo 21 eslabones, estadísticamente entre 1 millón se puede esperar por lo menos una secuencia). Ácidos nucleicos “enantiomeras puras”, pueden, como se mostró atar prontamente, otras bases nucleótidos complementarios a la cadena del ARN, y construir helicoides dobles bastante estables. Por fluctuaciones del medio ambiente, estos entonces pudieron, durante los ciclos de temperaturas constantes durante el día cálido, disociarse en cadenas individuales, que en las noches frescas re-combinarse con bloques de ADN ópticamente estrógenos y así sucesivamente (también esto esta estadísticamente documentado, Kämpfe 1992). Mediante procesos de este tipo (como adecuado, hoy también se consideran los entorno calientes de los “black smokers” de las profundidades marinas) se posibilitó la reproducción libre de Replicasa (Duplicación) durante el ritmo circadiano, donde las copias del ADN pudieron juntarse a cuasi-especies o estructuras terciarias, donde actuaban, la velocidad de replicación, la exactitud del copiado y eventualmente propiedades catalíticas, termodinámicas y estéricas actuaban como factores selectivos. Con esto, después de la fase divergente de la, organización, tenemos por primera vez una evolución convergente de Darwin, la selección lleva automáticamente a una construcción de información y neguentropía. También la simetría óptica de las unidades de Ribosa, se puede explicar ineludiblemente mediante este modelo, debido a que sólo Oligómeros de ARN pueden uniformes, pueden garantizar la formación de doble hélices estables, con exactitud copiadoras y altamente replicativos.

En una superficie (a) (cadena colectora) se pueden atar moléculas de fijación (b) (por ejemplo cadenas abiertas Oligonucleótidas), que por su lado, muestran una fuerte afinidad hacia los aminoácidos (c). Todo este arreglo puede, como cree Wächterhäuser, adherir o formar sobre una superficie de cristales de pirita, arcillas, arena o lava, , donde la pirita como también otros sulfuros metálicos tienen propiedades catalíticas favorables, por ejemplo para la formación de proteínas que realizaron el enlace de los aminoácidos a una cadena de proteínas primariamente estructurada; se formó el aparato traductor originario, Con el correr del tiempo se formaron los adecuados híper-ciclos de encima-ácido nucleíco, que formaron un sistema auto-replicante y auto catalizador.
Ahora se sabe que ácidos nucleicos se pueden unir con estructuras terciarias para formar una especie de ovillos, donde este tipo de unidades, bajo determinadas condiciones son capaces de atar a numerosos oligonucleótidos de forma de agujas que “sobresalen como alfileres de un alfiletero”. A estos oligonucleótidos rectilíneos, Kuhn los llama “filamentos colectores” los que, según lo sugerido por nuestras actuales conocimientos, habrían sido capaces, nuevamente por su lado atar, por intermedio de puentes de Hidrógeno cortas secuencias de poli nucleótidos complementarios (“Moléculas de fijación”). Las moléculas de fijación según Kuhn, formaron los t-ARN originarios, los filamentos colectores de los m-ARN originarios. Estos complementos bastante complejos pueden adicionar aminoácidos en las moléculas de fijación, entonces la formación de proteínas y encimas complejas, es relativamente posible. Junto a estos complementos, según Katchalsky también pueden venir al caso sustancias trazadoras inorgánicas con propiedades catalíticas, como la pirita y montmorillonita. Como puede verse en las figuras 12 y 13, a partir de esto, se forma el aparato traductor original, debido a que los aminoácidos fueron ligados a las moléculas de fijación y que por la catálisis de las superficies de los agregados, aquellos fueron transformados en encimas. El puente hacia la teoría del metabolismo de superficie de Wächterhäuser es evidente.
b) La teoría de los aciertos múltiples
En oposición a la hipótesis de los pasos múltiples de Kuhn, Kaplan postula una formación de los primeros protobiontes en un sólo paso, donde azarosamente sintetasas y ácidos nucleicos deben haberse encontrado en un compartimiento. De cada una de las dos unidades debió haber existido un “repertorio básico” de macro moléculas, que se organizaron auto catalíticamente en asociaciones. Kaplan estimó las condiciones marcos y después de consideraciones estadísticas, llegó a la conclusión, que la formación de sistemas auto catalíticos pueden haber estado del todo en el campo de las posibilidades.-
Bibliografía
Holleman, Wiberg: Lehrbuch der Anorganischen Chemie, S. 518 f. Berlin, 1995
E. W. Bauer et al: Biologiekolleg. Bielefeld, 1983
Bilder frühen Lebens. Verständliche Forschung, Spektrum der Wissenschaft. Heidelberg, 1986
Entwicklung von den ersten Lebensspuren bis z. Menschen. Verständl. Forschung, Spektrum d. Wiss.. Heidelberg, 1988
Darwin, Charles: Die Entstehung der Arten durch natürliche Zuchtwahl. Stuttgart, 1963
Hoimar v. Ditfurth: Am Anfang war der Wasserstoff. Hoffman und Campe, 1980
Stuart Kauffman: Der Öltropfen im Wasser, 1995
L. Kämpfe: Evolution und Stammesgeschichte der Organismen, editorial Gustav-Fischer, 1992
Traducido del alemán por A. Gundelach, Marzo 2012, con la gentil autorización del Dr. Martin Neukamm "Scritte zum leben"

















No hay comentarios.:
Publicar un comentario