La evolución en la mira del creacionismo
1.- Bases científicas teóricas
Clasificación de las aéreas de conocimiento
Clasificación de las aéreas de conocimiento
En el sentido más estricto de las ciencias reales, son ciencia, aunque la tecnología, las matemáticas y las ciencias espirituales igualmente producen conocimientos fiables y están estrechamente ligadas a las ciencias reales. Frente a estas, las tal llamadas para-ciencias solo producen conocimientos ilusorios.
Ciencias reales
|
Ciencias
|
||||
Técnica
|
Tecnología
|
Matemáticas
|
Ciencias espirituales
|
Conocimientos cotidianos
|
Ciencias confiables
|
Para-tecnología
|
Para-ciencias
|
Ciencias para-espirituales
|
No son ciencias
|
2.- La evolución como blanco de la crítica teórica científica
Esquema de una explicacion evolucionaria
Premisa 1:
Los genes son heredados a los descendientes, es decir de la especie de los antepasados a la especie de los descendientes (Postulado de la descendencia)
Premisa 2:
Nuevos genes se originan, entre otras cosas, por diferentes mecanismos de la mutación de genes y cromosomas (por eje. duplicación)
• Genes funcionalmente diferentes de un organismo muestran muchas similitudes secuenciales
• Algunos genes repiten, en parte muchas veces las mismas secuencias de ADN
La teoría de la descendencia de Darwin atada a los conocimientos adicionales sobre los mecanismos de la herencia, la duplicación y mutación de los genes, e implícitamente la teoría de la selección forman el “marco” explicativo, de los cuales se pueden sacar conclusiones concretas que se pueden comprobar y validar empíricamente
Explicación mediante el diseño inteligente
Premisa 1:
Los genes son heredados a los descendientes, es decir de la especie de los antepasados a la especie de los descendientes (Postulado de la descendencia)
Premisa 2:
se forman nuevos genes por un diseño inteligente.
Conclusión:
• Los genes en su secuencia se asemejan entre sí o son totalmente únicos ???
• Los genes son repetitivos o no repetitivos ???
Ya que en el “Designer” se trata de un proceso totalmente desconocido y de factores totalmente imposibles de investigar, no es posible dar límites objetivos para la acción de un diseñador. Luego no es una explicación específica ni causal, y tampoco existe la posibilidad de una comprobación a mano de conclusiones concretas. Los anti evolucionistas con sus explicaciones sacan a las explicaciones científicas de sus bases y por lo tanto caen detrás del estado de la teoría de la evolución.
3.- Evolución – sin casualidad. La argumentación con la probabilidad
Evolución del ojo lenticular

Células epitetiales
Fibras verviosas
c Cavidad con líquido
d-e Córne
Lente
Nervio óptico
En primitivos poli celulares, pigmentos especializados permiten una percepción de oscuridad y claridad (a), donde algunos invertebrados se encuentran dentro de unas hendiduras protectoras. Esta función protectora puede ser valorada positivamente en cada paso. A partir de una determinada profundidad surge con toda naturalidad otra ventaja (doble función), a ser una simple “visión direccional y de movimiento”: se formó el “ojo de copa” (b). Mientras más estrecha es la abertura para la entrada de la luz, tanto más nítida es la vista direccional, de manera que paso a paso bajo un constante control selectivo se puede formar un “ojo cámara oscura” (c). Nuevamente fue una función doble, añadido al hecho, que adicionalmente se formaba un órgano. El cierre del ojo por una piel translucida y la aparición de una sustancia gelatinosa nuevamente favorecen por razones de protección. Como la imagen ahora es nítida pero con una luminosidad débil, se da la posibilidad de una mejoría de las propiedades ópticas del ojo, en especial para la formación de una lente convergente, que una nítida con luminosidad, d: el “ojo vesicular”, e: el ojo de los vertebradosEn primitivos poli celulares, pigmentos especializados permiten una percepción de oscuridad y claridad (a), donde algunos invertebrados se encuentran dentro de unas hendiduras protectoras. Esta función protectora puede ser valorada positivamente en cada paso. A partir de una determinada profundidad surge con toda naturalidad otra ventaja (doble función), a ser una simple “visión direccional y de movimiento”: se formó el “ojo de copa” (b). Mientras más estrecha es la abertura para la entrada de la luz, tanto más nítida es la vista direccional, de manera que paso a paso bajo un constante control selectivo se puede formar un “ojo cámara oscura” (c). Nuevamente fue una función doble, añadido al hecho, que adicionalmente se formaba un órgano. El cierre del ojo por una piel translucida y la aparición de una sustancia gelatinosa nuevamente favorecen por razones de protección. Como la imagen ahora es nítida pero con una luminosidad débil, se da la posibilidad de una mejoría de las propiedades ópticas del ojo, en especial para la formación de una lente convergente, que una nítida con luminosidad, d: el “ojo vesicular”, e: el ojo de los vertebrados.
Formación de la región T-urf13 en el genoma del maíz de la variedad CMS-T

Arriba en el cuadro está representado el gen 26s rARN mitocondrial del maíz, bien abajo el T-urf13 (con los codones de partida y de detención ATG, TGA). Sobre el gen T-urf13 están representados los dos fragmentos de gen que se fundieron hacia el gen T-urf13; ambos fragmentos se pueden derivar de la misma cuerda del 26s rARN. El signo interrogatorio simboliza aquella parte del gen, que presumiblemente se formó por la inserción de optros fragmentos de genes. Modificado según Hunt (2007).
Estructura del sistema de flagelos de la Escherichia coli
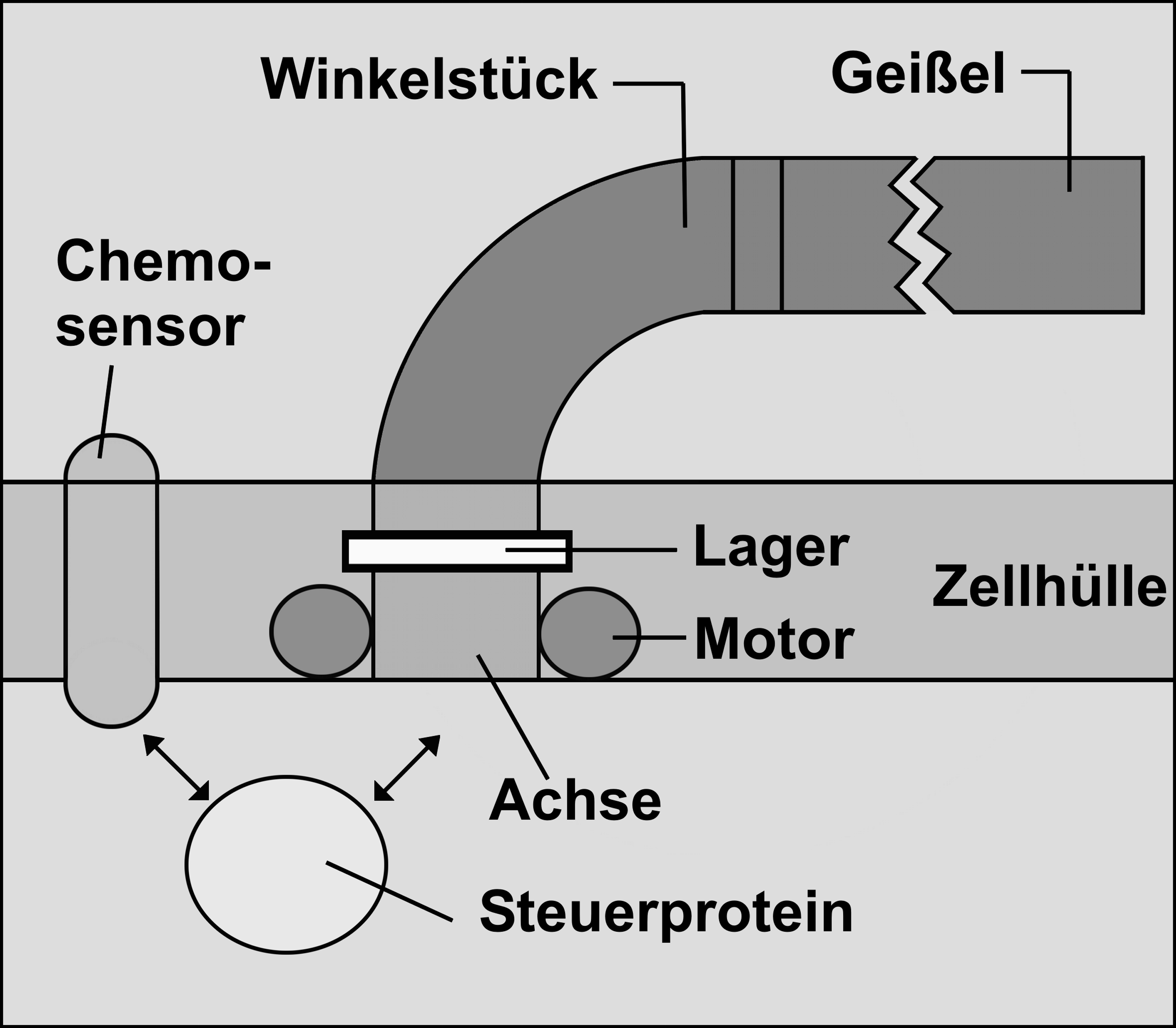
Winkelstpúck = Codo
Geißel = Flagelo
Lager = Cojinete
Zellhülle = Membrana celular
Achse = Eje
Steuerprotein = proteína de mando
El sistema del flagelo, según Junker/Scherer (2006) se compone de 7 elementos básicos: Un flagelo rotatorio; un codo; una es0pecie de sensor químico; un cojinete; del “motor bacterial”, el eje y una proteína de mando. Ya que ahora todos estos elementos básicos son necesarios, el sistema del flagelo es irreduciblemente complejo: Si se retira sólo uno de los componentes el sistema de tracción rotatorio ni funciona. Consecuentemente todo este sistema supuestamente debe haber sido construido en un solo paso, para ser favorecido por primera vez por la selección. Esta afirmación, después de una mirada más de cerca resulta ser falsa (ver el siguiente cuadro). Modificado según Junker/Scherer
Etapas de la evolución del sistema del flagelo
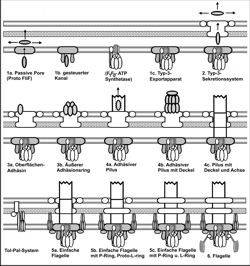
Un poro pasivo en el interior de la membrana celular puede servir como punto de partida, por una adición posterior de una ATPasa se transformó en un primitivo sistema de secreción bacteriano Tipo III. Con la adición de una tal llamada secretina en el siguiente desarrollo, también en la membrana celular exterior se forma un poro, de manera que pueden ser transportado sustancias hacia el entorno de la célula, tal vez para traspasar sustancia nutritivas presentes en una forma transportable para el organismo. Con el correr del desarrollo se presenta ahora la opción, mediante “proteínas de adhesión” que se adhieren en el anillo exterior del sistema de secreción, usan el potencial para la adhesión de sustratos favorables para el organismo (“función de anclaje”). Diferenciaciones en forma de pili ahora son posibles en forma continua. Si la función de anclaje de los pilus no es aprovechado, entonces estos también pueden servir para transmisión de la fuerza del momento angular de las motor-proteínas al medio circundante. Probablemente para esto bastaría el reclutamiento de las proteínas TolQR y ExbBD las cuales con el aprovechamiento de un gradiente de protones pon e a rotar al pilus. Una flexión optimiza la propulsión (ventaja selectiva: Movilidad). . Ya que la ATPasa y las proteínas TolQR y ExbBD se encuentran en el lugar “correcto” en la membrana y la mayoría de proteínas, necesarias para el motor bacteriano, ya se encuentran en el organismo, sólo el armado gradual de los componentes. Modificado según Matzke (2003) .
La reconstrucción de la filogenia
Cladograma de los amniotas (propietarios de un envoltorio embrionario)

Amnioten = amniotas
Sauropsiden = Saurópsidos
Diapsiden = Diápsidos
Eidechsen = Lagartos
Kokodrile = Cocodrilos
Vögel = Aves
Wirbelsäule = Columna vertebral
Embryonhülle = Envoltorio embrionario
Hautschuppen = Escamas
Zwei Temporalfenster = Dos aberturas del hueso temporal
Unterkieferloch, Präorbitalfenster = Orificio en la mandíbula inferior , abertura pre orbital
Schnabel, Federn = Pico, plumas.
De acuerdo al cladograma, las aves y los cocodrilos provienen de una de una clase de ancestro común (especie parental). Grupos como estos, que provienen de una ramificación se les llama también grupos hermanados. El taxón de los arcosaurios reúne a las aves y los cocodrilos en un nuevo grupo. Este grupo, por su lado, es el grupo hermanado de los lagartos, esto significa, que sus representantes provienen de una rama genética común. El grupo de los arcosaurios y los lagartos por su lado son agrupados en el taxón de los diápsidos, que de nuevo es el grupo hermanado de las tortugas, etc.
Evolución de la articulación secundaria de la mandíbula

dos articulaciones mandibularesm(por eje. el Diarthrognathus. Después el angulare inicialmente se achicó, con el tiempo se separó de la mandíbula inferior transformándose en el hueso timpánico (Ty) que soporta al tímpano. Al mismo tiempo se produjo el desprendimiento del articulare de la mandíbula inferior, como el quadratum del cráneo y su desplazamiento hacia el oído medio – formando el martillo y el yunque. Modificado según Theobald (2004) y Kargong (2002)
(früher Säuger = Mampifero antiguo)
¿Los huesos del oído medio se originaron en forma convergente?

* Formación de tres huesecillos del oido interno
Citando un dudoso artículo (Rich et al. 2005) la asociación creacionista “Wort und Wissen”, el fósil del monotremata /Teinolophos/ no permite otra interpretación desde el punto de vista de la evolución, que “ya en si mismo, la improbable transformación de la articulación tamporomandibular ósea”, del mamífero se llevó a cabo de forma independiente, por lo menos dos veces. De ello se desprende la pregunta retórica, en qué medida esta situación, desde el punto de vista de la teoría de la evolución, esto aun “se puede interpretar de manera plausible”. Estudios más recientes, empero refutan la hipótesis de la formación convergente de los huesecillos del oído medio (Rowe et al. 2008).
Arriba: Hipótesis de parentesco según Rich et al. (2005). Según esto el origen de los tres huesecillos fue convergente.
Abajo. El esquema revisado según Rowe et.al (2008). Las estrellitas, que representan la formación de los huesecillos del oído, se juntan ahora en un solo punto.
Simulación de Monte-Carlo de la dinámica evolutiva

Ïndice de diversidad (por eje. cantidad de especies)
Tiempo (generaciones)
Rápidos impulsos de desarrollo (Exterminación masiva o veloz diversificación)
Cuasi estado estacionario
En el proceso numérico se consideraron valores conocidos, como el crecimiento de poblaciones, mutación, selección, formación de especies, y exterminios y descritos mediante funciones matemáticas. Bajo la consideración de la ley “de la gran cifra” [ley de Benford (el traductor)] se confirma en el resultado del cuadro del postulado de la teoría de la evolución: Largas fases de estasis son interrumpidos por períodos de altas fluctuaciones en vista de la diversidad de las especies. Como en la simulación sólo entraron antecedentes conocidos, el resuktyado se ouede considerar como una confirmación independiente del modelo puntual de la evolución. Según Rikvold/Zia (2003)
¿El Archaeoopoteryx –Reptil o ave?

Los esqueletos en comparación del Coleosaurio, del Archaeopteryx y de una ave moderna (una paloma). P: Pigostilo, G: Gabelbein: Fémur (?, el trad.), Brustbein: Esternón. Compsognathus según una plantilla de Matthias Kabel (de.wikipedia.org/wiki/Compsognathus). Archaeopteryx modificado según Bruno Kremer (1986): El ave originaria Archaeopteryx, esqueleto de una paloma según una fotografía de Uwe Gille (de.wikipedia.org/wiki/Vogelskelett).
La evolución de los tetrápodos

La conquista de la tierra firma por parte de los tetrápodos originarios hace aprox. unos 380 a 350 millones de años en el Devónico. La transición de la vida acuática hacia una vida en tierra firme se puede visualizar a manos de las articulaciones de los huesos de las aletas pectorales del Eusthenopyeron y Panderichthys como también de los tetrápodos Ichthyostega y Tulerpeton del Devónico. Las articulaciones de las alteas pectorales del Tiktaalik continúan con muchas características individuales. Según Shubin et al. (2006).
Biología evolutiva del desarrollo (“evo-devo”)
Los genes homeóticos en la mosca de la fruta y ratones

Fliege = mosca
Maus = ratón
Organización de los genes homeóticos de la clase Hom/Hox y su conservación
en la evolución. Los genes homeóticos
están localizados en dos grandes complejos genéticos que son llamados complejo
Antennapedia y complejo Bithorax (Ant-C
y BX-C). Los miembros numerados de un complejo muestran una alta correlación en
su consecuencia. Lo que se puede explicar con una repetida duplicación de los
genes homeobox en el correr de la evolución. En la rata estos complejos genéticos
aparecen cuatro veces, de manera que probablemente hubo dos duplicaciones de unos
antecesores comunes de insectos y mamíferos. Números iguales simbolizan homeo
genes con una especial alta concordancia de secuencias. Las enumeraciones en el
embrión del ratón (abajo), indican en que zonas del cuerpo controlan los genes
hox, la formación de l,os órganos. Según
Müller/Hassel (2005).
Esquema genético del desarrollo de la "caja de herramientas"

Esquema genético del desarrollo de la "caja de herramientas"
Mastergene.por eje. Pax6-, dII, genes Tinman etc.
Transposones, Gentinquerin
Reguladores del tipo de células
Proteínas señalizadores
Receptores de células.
Hormonas
Esquema del desarrollo genético de la “caja de herramientas”. Mutaciones en la vía germinal pueden influenciar la regulación de genes en el desarrollo embrionario. Si por ejemplo los “genes maestros” son afectados por las mutaciones o genes que regulan el vaciado de hormonas, entonces los genes objetivos son activados en forma distinta espacial y temporalmente desactivados o determinadas células funcionalmente alteradas. La expresión de genes alterados, en el desarrollo embrionario puede causar múltiples cambios en el fenotipo. De esta manera con el correr del tiempo se pueden formar rasgos complejos. Según Carroll (2008).
La aparición de estructuras complejas por tinkering genético
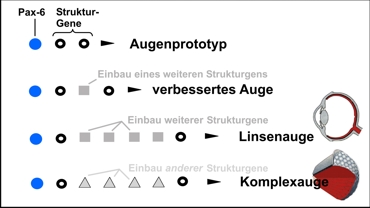
Einbau eines weiteren Strukturgens = Incorporación de otro gen estructural
Verbessertes Auge = Ojo mejorado
Einbau weiterer Strukturgene = Incorporación de otros genes estructurales
Linsen ague = ojo lenticular
Komplexauge = Ojo complejo
Representación esquemática del tinkering genético. Su la evolución comienza con un conmutador principal (por eje. Pax-6) como también con algunos genes estructurales intermedios, bajo un continuo control selectivo en forma sucesiva otros genes pueden ser puestos bajo en control de este gen maestro, con lo cual la cascada de desarrollo se hace cada vez más compleja. De esta manera, por ejemplo de una mancha sensible a la luz (ojo prototipo), se formó paso a paso a paso un ojo lenticular y el ojo complejo.Según Junker/Scherer (2006), de acuerdo a Gehring (2001).
La diferenciación funcional de las redes de genes
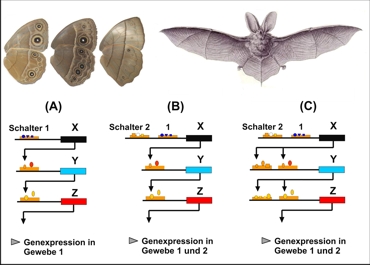
Schalter = Conmutador
Gene Expression in Gewebe = Expresión genética en tejidos
Arriba: El gen regulador dII (dlx), participa en el desarrollo de estructuras tan diferenciados como los patrones de las alas de las mariposas y las extremudades de los mamíferos (por eje. el murciélago).
Abajo: (A) esquema de una malla de genes (X,Y, Z,) simple. El gen X controla la expresi’p0on del gen Y, el cual, en forma análoga regula la expresión de Z. La secuencia reguladora (“conmutador 1”), delante del gen X, se encarga que se activa esta cascada en el tejido 1. (B) si ahora un segundo conmutador, que puede ser activado en el tejido 2, es ub9cado delante del gen X (“Conmutador 2”), entonces esta cascada también se hace activa en este tejido. Se originó una función dual. (C). Modificaciones posteriores de uno o varios conmutadores facilitan diferenciaciones funcionales escalonadas, de manera que esta cascada en distintos tejidos puede cumplir con funciones específicas respectivamente. Según Monteiro/Podlaha (2009).
Árbol genealógico comparativo de 337 de single-copy genes
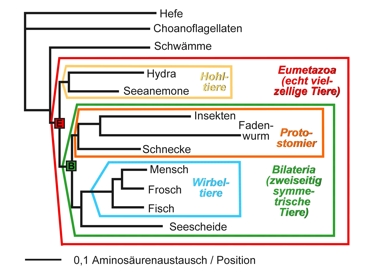
Choanoflagellaten = Coanoflagelados
Schwämme = Esponjas
Seeanemne = Anémonas marinas
Echt vielzellige Tiere = Auténticos poli celulares
Hohl-tiere = Celenterados
Insekten = Insectos
Faden-wurm = nematodo
Schnecke = Caracol
Proto-stomier = Protostomia
Mensch = Humano
Frosch = Sapo
Seescheide = Ascidia
Wirbekltiere = Animales vertebrados
Zweiseitig symetrische Tiere = Animales bisimétricos
0,1 Intercambio de aminoácidos/posición
Se compararon los genes de la Ascidia (Ciona), pez (Takifugu), sapo (Xenopus), ser humano, caracolm(Lottia), mosca (Drosophila), nematodo (Caenorhabditis), pólipo de agua dulce (Hydra), anénomas marinas (Nematostella), esponja (Amphimedon), coanoflagelados (Monosigha) y levadura (Saccharomyces). Todos los puntos de ramificación es pudieron ser resueltos con una fiabilidad del 100%. Escala: 10% de probabilidad de intercambio. E: El ancestro de todos los eumetazoas. Hasta llegar a él se formaron 1148 nuevos genes y 1470 genes por duplicación. La cantidad de sus genes que pudieron ser reconstruidos es de 7766. B: Ancestro de todos los bilanterians,. Hasta él (es decir desde ”E”) se originaron 662 nuevos genes y 320 genes por duplicación de genes. Según Putnam et. al (2007).
Información general sobre el origen de la reserva genética en animales
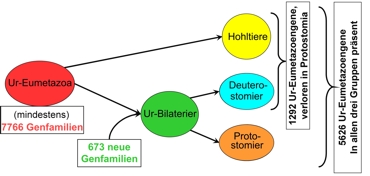
Izquierda: Repertorio genético de los E, como a la derecha: donde los Celenterados y deuterostomes. Para cada derivación, fue posible dar cifras en referencia a las pérdidas y formación de genes; para mayor claridad aquí sólo se mencionan unos pocos. A esto se le agregan nuevas formaciones por duplicación de genes (obviados aquí). Según los cálculos de los autores, por eje. se pueden derivar 13830 genes humanos por este mecanismo, del reserva originaria de los eumetazoas. Según Putnam et al, (2007).
La evolución química
Representación de la quiralidad
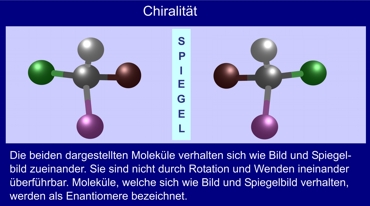
Las dos moléculas representadas, se comportan entre sí como imagen e imagen reflejada. No son transferibles entre si por rotación o volteo, aquellos que se comportan como imagen e imagen refglejada son llamados enantioméricos
Asimetría o quiralidad respectivamente en base de cuatro diferentes grupos atómicos en un átomo de Carbono. Constelaciones como estas pueden aparecer varias veces en grandes moléculas. En apoyo a Michael Rainer A. Müller (chempage.de/lexi/chiral.htm).
Cuotas de formación de homoquirales de proteínas auto replicantes

Produktmenge = Cantidad de producto
Zeit = Tiempo
Zeit = Tiempo
El laboratorio de Reza Ghadiri del Scripps Research Institute en La Jolla (California), en el año 1996 descubrió un poli péptido auto replicante (Lee et al. 1996). ¡Este poli péptido, que se condensado de 32 aminoácidos, sirve como matriz y apoya su propia generación auto catalítica! Lo especial en esto es: Tiene la propiedad, que en su replicación en un proceso de selección prefiere formar productos homoquirales. Estos productos nuevamente sirven como matrices, que nuevamente originan productos de una mayor pureza enantionmérica y así sucesivamente. Con esto disminuye la capacidad catalítica, si sólo un componente muestra una lateralidad opuesta a uno de los demás aminoácidos. Es lógico que un mecanismo como este ha influenciado la formación de la homoquiralidad en la Tierra originaria. La gráfica muestra las cuotas de formación de los productos en dependencia del tiempo. La generación dependiente de las matrices: a) de los productos homoquirales.b) de los productos heteroquirales. Según Saghatelian et al. (2001).
El concepto del Chemoton
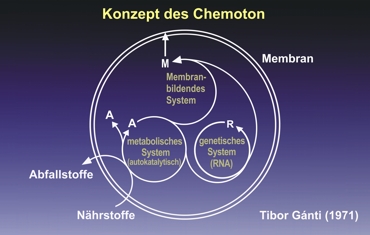
M: Membran = Membrana
M: Menbran bildenes System = Sistema formador de membranas
A: Metabolisches System (autokatalitisch) = Sistema metabólico (auto catalítico)
R: Genetisches System (RNA) = Sistema genpetico (ARN)
A: Abfallstoffe = Sustancias de deshecho
Nährstoffe = Sustancias nutritivas
M: Menbran bildenes System = Sistema formador de membranas
A: Metabolisches System (autokatalitisch) = Sistema metabólico (auto catalítico)
R: Genetisches System (RNA) = Sistema genpetico (ARN)
A: Abfallstoffe = Sustancias de deshecho
Nährstoffe = Sustancias nutritivas
Un modelo muy interesante es el muy citado “Chemoton” del científico húngaro T. Gánti. En la bibliografía relevante es considerado como modelo del sistema más pequeño auto replicante ("minimal unit of life"). Según esto están acoplados tres subsistemas que se auto reproducen. Este acoplamiento lleva a la proliferación al y controlado programáticamente, “fluyente autómata”, el “Chemoton”. Es mucho menos complicado que varios esquemas complejos excesivamente exagerados de un metabolismo mínimo presentado por los anti evolucionistas. Ilustración según Günther von Kiedrowski; con la gentil autorización del autor.
La producción de proteínas en la célula
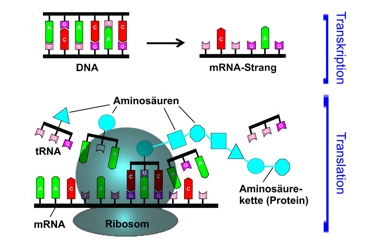
En primer lugar, a partir del gen del ADN, que codifica para la proteína, instalado una copia en forma de una cuerda de ARN (Transcripción). La información es usada entonces con el correr de la translación, para la manufactura de la proteína. Con ello codifican cada vez tres bases nucleícas vecinas sobre el ARNm (Codons/base triplete) un determinado aminoácido, del cual la proteína es construida paso a paso., Como transportador funciona el tal llamado ARNt (Transfer-ARN). Este está cargado con un aminoácido y con uno de sus extremos (el llamado anti codón) puede acoplarse exactamente en una base triplete en un ARNm. El ribosoma junta de tal manera al mARN y tARN cargado, de tal manera que la base triplete del mARN y el correspondiente anticodón (complementario) del tARN, se adhieren.Los aminoácidos de dos tARN vecinos son entonces unidos, y el tARN abandona entonces el ribosoma sin en aminoácido. Entonces se acopla la siguiente apropiada molécula tARN apropiada al mARN, con lo cual el aminoácido correspondiente es atado a la ya existente cadena de aminoácidos. Este proceso continúa hasta que un codón de detención interrumpe el proceso y señaliza que la cadena de aminoácidos está completa.
Crítica a la "macro evolución": Complejidad irreducible
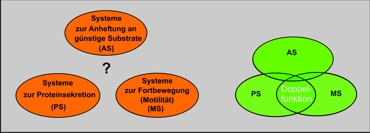
Systeme zur Anheftung an günstige Substrate (AS)
Sistemas para la adhesión a sustratos favorables
Sistemas para la adhesión a sustratos favorables
Systeme zur Proteínsekretion (PS)
Sistemas para la secreción de proteínas
Systeme zur Fortbewegung (Motilität) (MS)
Sistemas para la locomoción (motricidad)
Izquierda: Como se imaginan la evolución los anti evolucionistas. Las elipses representan tres cantidades de sistemas de bacterias, que cumplen, cada cual con determinadas funciones. El campo entre las tres cantidades (estados de funcionamientos básicos), está vacío, esto significa, que presuntamente no puede ser subdividido en etapas selectivamente positivas (Junker/Scherer 2006, 158). En consecuencia se requiere una gran acumulación de mutaciones adecuadas, para llegar un estado funcional al siguiente, lo que sería muy poco probable (“problema de construcción”). Dentro de las elipses, sin embargo se concede una micro evolución (”problema de optimización”).
Derecha: Como la moderna biología se imagina la evolución: Los atributos, según el estado pueden ejercer no sólo una función, sino varias al mismo tiempo (multifuncionalidad). Se producen súper posiciones funcionales, con lo cual se desarrolla un camino revolucionario viable.
Evolución de los sistemas irreductiblemente complejos

Estructura X con función a
Estructura X + elemento A → mejoramientode la función a
Estructura XA + elemento B → otro mejoramiento de la funciónm
Estructura XAB + elemento C → nueva, adicional funcipon b
Estructura XABC + elemento D → la nueva función b es mejorada
El elemento b es modificado → Los demás elementos se hacen indispenzables para la función b
Formación gradual de un sistema irreductiblemente complejo (sistema IC) por optimización de una vía lateral: La función a de una estructura X, primeramente por la adición de los elementos de construcción A, B y C es gradualmente optimizado. La nueva formada estructura XABC, realiza ahora súbitamente una total nueva (adicional) función b. En consecuencia de nuevos pasos optimizadores (adición de un elemento D y las modificaciones de los elementos que ya existen) se establece la función b paso a paso en el sistema. Partes redundantes son eliminados u modificados. De esta manera todos los elementos se hacen indispensables para la función. Dibujado con referencia a Orr (1996).
Lo que supuestamente la selección no puede realizar
Esquema dela evolución de plantas carnívoras


Inflorescencia de la Araceae (3)

Lo que supuestamente la selección no puede realizar
Esquema dela evolución de plantas carnívoras
Espacio
vital
|
Trampas pegajosas
y de bisagras
|
Formas de
transición
|
Trampas deslizadoras
y derivadas
|
Terrenales,
Trampa en
el aire
|
Delgadas hojas
captoras
Triphyophylum,
Drosophylum,
Drosera regia
Byblis
|
Recipientes tubulares
Con paredes interiores
Pegajosas
N. infundibuliformis
|
Trampas de fosas
tubulares erectas |
Terrestres
nivel suelo |
Anchas hojas
Atrapadoras
Pinguicula vulgaris
Drosera
schizandra
|
Fuertes hojas enrolladas
Atrapadoras
Pinguicula
lutea
|
Jarras cortas
recogidas
|
Anfibio
|
Trampas con un
mecanismo de
cierre rápido
|
Trampa de deslizamiento
y tipo red
|
|
Zonas de
inundacioines
|
Trampas tipo
Bisagras
acuáticas
|
Trampas
acuáticas
de redes y succión
|
Inflorescencia de la Araceae (Arum)
Izquierda: Orontium aquaticum, espádice sin espata adyacente, flores hermafroditas con perianto.
Derecha: Spathiphyllum wallisii. Espata similar a hoja de follaje con nervio central, después cambiando a verde, flores hermafroditas con perianto.
Derecha: Spathiphyllum wallisii. Espata similar a hoja de follaje con nervio central, después cambiando a verde, flores hermafroditas con perianto.
Inflorescencia de la Araceae (2)
Izquierda: Anthurium waroqueanum. Espata discreta, flores hermafroditas, con perianto (discreto).
Derecha: Anthurium-andreanum-Hybride. Espata notoria, flores hermafroditas, con perianto (discreto)
Derecha: Anthurium-andreanum-Hybride. Espata notoria, flores hermafroditas, con perianto (discreto)
Inflorescencia de la Araceae (3)
Izquierda: Callopsis volkensii, espádice zonificada, parte femenina con espata.
Derecha: Lagenandra lancifolia, Trampa altamente compleja con función tipo snorkel
Traducido del alemán, por A. Gundelach, con la gentil autorización del Dr. Martin Neukamm. Agosto 2012


















{kind=link}
4 comentarios:
¿Es verdaderamente científica la enseñanza de la evolución?
El “método científico” es el siguiente: Observe lo que sucede; fundándose en esas observaciones, formule una teoría en cuanto a lo que posiblemente sea verdad; someta a prueba la teoría mediante observaciones adicionales y por experimentos; y vigile para ver si se cumplen las predicciones fundadas en la teoría. ¿Es este el método que han seguido los que creen en la evolución y la enseñan?
El astrónomo Robert Jastrow dice: “Para desazón suya, [los científicos] no tienen respuesta clara, porque los químicos nunca han logrado reproducir los experimentos de la naturaleza sobre la creación de la vida desde materia inanimada. Los científicos no saben cómo sucedió eso” (The Enchanted Loom: Mind in the Universe [El telar encantado: mente en el universo], Nueva York, 1981, pág. 19).
El evolucionista Loren Eiseley reconoció lo siguiente: “Después de haber regañado al teólogo por confiar en mito y milagro, la ciencia se halló en la posición no envidiable de tener que crear una mitología propia: a saber, la suposición de que lo que tras de mucho esfuerzo no pudo ser probado que estuviera aconteciendo hoy, había, en realidad, tenido lugar en el pasado primitivo” (The Immense Journey [El larguísimo viaje], Nueva York, 1957, pág. 199).
Según la revista New Scientist: “Una cantidad creciente de científicos, y más particularmente un número aumentante de evolucionistas [...] presentan el argumento de que la teoría evolucionista darviniana no es de modo alguno una teoría genuinamente científica. [...] Muchos de los críticos tienen las más elevadas credenciales intelectuales” (25 de junio de 1981, pág. 828).
El físico H. S. Lipson dijo: “La única explicación aceptable es creación. Sé que esto es anatema para los físicos, como de hecho lo es para mí, pero no debemos rechazar una teoría que no nos gusta si la prueba experimental la apoya”. (Cursivas nuestras.) (Physics Bulletin, 1980, Vol. 31, pág. 138.)
¿Están en armonía entre sí los que apoyan la evolución? ¿Cómo afectan estos hechos lo que usted piensa en cuanto a lo que ellos enseñan?
La introducción a la edición centenaria de El Origen de las Especies, de Darwin (Londres, 1956), dice: “Como sabemos, hay gran divergencia de opinión entre los biólogos, no solo en cuanto a las causas de la evolución, sino hasta en cuanto al proceso mismo. Esta divergencia existe debido a que la prueba es insatisfactoria y no permite llegar a ninguna conclusión segura. Por lo tanto, es correcto y apropiado llamar la atención del público no científico a los desacuerdos que existen respecto a la evolución” (por W. R. Thompson, entonces director del Instituto de Control Biológico de la Comunidad Británica de Naciones, Ottawa, Canadá).
“Un siglo después de la muerte de Darwin, todavía no tenemos ni la más ligera idea demostrable, o siquiera plausible, de cómo en realidad tuvo lugar la evolución... y en los últimos años esto ha llevado a una serie extraordinaria de batallas en cuanto a toda la cuestión. [...] Entre los evolucionistas mismos existe un estado de casi guerra abierta, en la que toda clase de secta [evolucionista] insta a que se efectúe alguna nueva modificación.” (C. Booker, escritor del Times de Londres, The Star, Johannesburgo, 20 de abril de 1982, pág. 19.)
La revista científica Discover dijo: “La evolución [...] no se halla solo bajo ataque por cristianos fundamentalistas, sino que también está siendo cuestionada por científicos de reputación. Entre los paleontólogos, científicos que estudian el registro fósil, aumenta la disensión” (número de octubre de 1980, pág. 88).
¿Qué punto de vista tiene el apoyo del registro fósil?
Darwin reconoció: “Si numerosas especies [...] en realidad han comenzado su existencia de una vez, ese hecho sería mortal para la teoría de la evolución” (The Origin of Species [El origen de las especies], Nueva York, 1902, Parte II, pág. 83). ¿Indica la prueba existente que “numerosas especies” llegaran a la existencia al mismo tiempo, o señala a desarrollo gradual, como afirma la enseñanza de la evolución?
¿Se han hallado suficientes fósiles como para llegar a una conclusión sólida?
Porter Kier, científico de la Institución Smithsoniana, dice: “En museos de todo el mundo hay cien millones de fósiles, todos catalogados e identificados” (revista New Scientist, 15 de enero de 1981, pág. 129). A Guide to Earth History (Guía a la historia de la Tierra) añade: “Con la ayuda de los fósiles los paleontólogos pueden darnos ahora un cuadro excelente de la vida de las edades pasadas” (Nueva York, 1956, Richard Carrington, edición Mentor, pág. 48).
¿Qué muestra, realmente, el registro fósil?
El Bulletin del Museo Field de Historia Natural de Chicago señaló: “La teoría de [evolución] de Darwin siempre ha estado estrechamente relacionada con prueba procedente de los fósiles, y probablemente la mayoría de la gente supone que los fósiles suministran una parte muy importante del argumento general que se presenta a favor de las interpretaciones darvinianas de la historia de la vida. Por desgracia, esto no es rigurosamente verdadero. [...] en aquel tiempo, tal como ahora, el registro geológico no presentó una cadena delicadamente graduada de una evolución lenta y progresiva” (enero de 1979, Vol. 50, núm. 1, págs. 22, 23).
A View of Life (Una vista de la vida) declara: “Comenzando a la base del período cámbrico, y extendiéndose por unos diez millones de años, todos los grupos principales de invertebrados ‘esqueletizados’ se presentaron por primera vez en el más espectacular aumento de diversidad que haya acontecido sobre nuestro planeta” (California, 1981, Salvador E. Luria, Stephen Jay Gould, Sam Singer, pág. 649).
El paleontólogo Alfred Romer escribió: “Debajo de esto [el período cámbrico], hay un vasto grosor de sedimentos en los cuales debería esperarse que estuvieran los progenitores de las formas cámbricas. Pero no los hallamos; estos lechos más antiguos están casi desprovistos de indicación de vida, y pudiera decirse que el cuadro general es consecuente, razonablemente, con la idea de una creación especial en el principio de los tiempos cámbricos” (revista Natural History, octubre de 1959, pág. 467).
El zoólogo Harold Coffin declara: “Si es correcto el concepto de una evolución progresiva desde lo sencillo hasta lo complejo, en el cámbrico se debería encontrar a los antecesores de estas criaturas vivientes totalmente desarrolladas; pero no se han hallado, y los científicos admiten que hay poca probabilidad de que alguna vez se hallen. Sobre la base de los hechos solamente, sobre la base de lo que en realidad se encuentra en la Tierra, la teoría de un súbito acto de creación en el cual fueron establecidas las formas principales de vida encaja mejor” (revista Liberty, septiembre/octubre de 1975, pág. 12).
Carl Sagan, en su libro Cosmos, reconoce con candidez: “La prueba fósil pudiera ser consecuente con la idea de un Gran Diseñador” (Nueva York, 1980, pág. 29).
¿Pudiera ser que el proceso evolutivo tuviera lugar como resultado de mutaciones, es decir, cambios drásticos que tuvieran lugar de súbito en los genes?
Science Digest declara: “Los revisionistas de la evolución creen que las mutaciones en genes regulativos clave pueden ser precisamente los ‘martillos neumáticos’ genéticos que su teoría de saltos significativos exige”. Sin embargo, esa revista también cita la siguiente declaración del zoólogo británico Colin Patterson: “El razonamiento superficial anda sin restricción. No sabemos nada acerca de estos genes maestros regulativos” (febrero de 1982, pág. 92). En otras palabras, no hay ninguna prueba en apoyo de la teoría.
The Encyclopedia Americana reconoce lo siguiente: “El hecho de que la mayoría de las mutaciones son dañinas al organismo parece difícil de conciliar con el punto de vista de que la mutación sea la fuente de materia prima para la evolución. Ciertamente los mutantes que se ilustran en los libros de texto de biología son una colección de fenómenos y monstruosidades, y la mutación parece ser un proceso destructivo, más bien que constructivo” (1977, tomo 10, pág. 742).
¿Qué hay de los “hombres-monos” en las ilustraciones de libros de texto, enciclopedias y museos?
“Para completar los detalles de la carne y el pelo de tales reconstrucciones hay que recurrir a la imaginación. [...] El color de la piel; el color, la forma y la distribución del pelo; la forma de los rasgos; y el aspecto de la cara... de estas características no sabemos absolutamente nada respecto a cualesquiera hombres prehistóricos.” (The Biology of Race, Nueva York, 1971, James C. King, págs. 135, 151.)
“La vasta mayoría de las concepciones artísticas se fundan más en la imaginación que en la prueba. [...] Los artistas tienen que crear algo que se encuentre entre un antropoide y un ser humano; mientras más antiguo se diga que es el espécimen, más parecido a mono lo hacen.” (Science Digest, abril de 1981, pág. 41.)
“Tal como lentamente estamos aprendiendo que los hombres primitivos no son necesariamente salvajes, así tenemos que aprender a darnos cuenta de que los hombres primitivos del período Glacial no eran ni bestias brutas ni semiantropoides ni cretinos. De ahí la inefable estupidez de todos los intentos por reconstruir al hombre de Neandertal o hasta al hombre de Pekín.” (Man, God and Magic [El hombre, Dios y la magia], Nueva York, 1961, Ivar Lissner, pág. 304.)
¿Acaso no se presenta la evolución como un hecho en los libros de texto?
“Muchos científicos sucumben a la tentación de ser dogmáticos, [...] vez tras vez se ha presentado la cuestión del origen de las especies como si finalmente se hubiera resuelto. Nada pudiera estar más lejos de la verdad. [...] Pero la tendencia a ser dogmáticos persiste, y no rinde ningún servicio a la causa de la ciencia.” (Periódico The Guardian, Londres, Inglaterra, 4 de diciembre de 1980, pág. 15.)
Pero ¿es razonable creer que todo lo que hay en esta Tierra fue creado en seis días?
Hay grupos religiosos que enseñan que todo lo que existe fue creado por Dios en seis días de 24 horas. Pero eso no es lo que la Biblia dice.
Génesis 1:3-31 da un relato de cómo Dios preparó la Tierra —que ya existía— para ser habitada por humanos. Dice que esto se efectuó durante un período de seis días, pero no dice que estos eran días de 24 horas. No es cosa rara en algunos países el que alguien se refiera a “el día de mi abuelo”, queriendo decir toda la vida del abuelo. Del mismo modo, la Biblia muchas veces usa el término “día” para describir un espacio de tiempo extenso. (Compárese con 2 Pedro 3:8.) Así, pues, razonablemente los “días” del capítulo 1 de Génesis pudieran haber durado miles de años.
Quiero aclarar algo. La evolución biológica no tiene nada que ver con el origen de la vida. La teoría de la evolución es un modelo explicativo de cómo ser formaron las distintas especias de un ser viviente originario. La evolución n o es una enseñanza, es parte integral de la biología
La teoría de la evolución de Darwin, fue reemplazada por la teoría sintética de la evolución. Hoy muchas cosas que Darwin hace mas de 150 años no sabía, desde entonces los conocimientos sobre el origen der las especies han aumentado enormemente, incluso desde 1981
No podemos basarnos en textos religiosos antiquísimos, para explicarnos el origen de las cosas
Las ciencias naturales no se basan en creencias ni en dogmas
NOTA.- Pronto voy a publicar un artículo que toma una posición científica referente a la pseudociencia del creacionismo
Publicar un comentario